
����ͨ��ʵ������ķ����������˲�ͬ��ɫ��ǿ���ѭ��ˮ��ֳϵͳ�������Ĵ���������������ʳ��Ӱ�졣
������ժҪ: ��������ʵ�鷨, �о��˲�ͬ��ɫ(��, A1; ����, A2; ���, A3)��������(24L�U0D, B1; 12L�U12D, B2; 8L�U16D, B3)��ǿ(0.88 W/m2, C1; 4.55 W/m2, C2; 8.60 W/m2, C3)��ѭ��ˮ��ֳϵͳ��������(850.97��82.77)g�Ĵ�������(Salmo salar)��������ʳ��Ӱ�졣ʵ����A1B1C1 (1)��A1B2C2 (2)��A1B3C3 (3)��A2B1C2 (4)��A2B2C3 (5)��A2B3C1(6)��A3B1C3 (7)��A3B2C1 (8)��A3B3C2 (9)9��������, ����Ӧ�趨����������180d���������, �ڹ�ɫΪ��⡢������Ϊ12L�U12D ��ǿ8.60 W/m2 �����´������ٵijɻ������, ����ɫ�������ں�ǿ�Գɻ��ʵ�Ӱ����첻����(P>0.05); ʵ���ڼ���������������ʺͷ����Ȳ����������(P>0.05); ����120 ��, 2��5��6 ������峤�ض��������������� 1 ��(P<0.05); ���� 180 ��, 1��2��4��7��8������������ض���������������6��(P<0.05), 1��2��3��4��7��8��9 �������������������6��(P<0.05), 9���������������ϵ����������7��(P<0.05)��9����Ѫ��������������������1��2��3��4��6��7 ��8 ��(P<0.05); ��ʳ�ʡ�����ת��Ч�ʺ�����ϵ�����ʱ�Ĺ�������Ϊ: ��⡢12L�U12D��8.60 W/m2, ����ɫ�������ں�ǿ����ʳ�ʡ�����ת��Ч�ʼ�����ϵ����Ӱ����첻����(P>0.05)����ʵ��������, ��Ϊ���˵Ĺ���������: ��⡢12L: 12D��8.60 W/m2��
������������(Salmo salar), �׳�������, ԭ���ڴ��������´����DZ�������, ����������Ҫ����ֳ����֮һ, ���й㷺�Ĺ������г�, ������Ϊһ��Ʒ���������г�ǰ���õ���ֳ�����������ڡ���������������ˮ������,�ҹ������غ������ļ�ˮ�½ϸ�,�����˺�ˮ������ֳ����ҵ��ѭ��ˮ��ֳϵͳ(Recirculating AquacultureSystem, RAS)���н�ˮ���������߲������ܵ�����������Ƶ��ص�, �ܹ�Ϊ��ֳ�����ṩ�ȶ��ɿص������������������������������з���,����ѭ��ˮ��ֳϵͳ�д��������ض�������ƫ��(0.33%~1.09%), ����ϵ���ϸ�(1.30~2.50), ����Ӱ���˲�ҵ�ķ�չ, ��˿�չѭ��ˮ��ֳģʽ�´���������������ʳ���ɵ��о�ʮ�ֱ�Ҫ���о�����, ���������Ӱ������������ʳ����Ҫ���ӡ�Head ��[1]�о�����, ��ͬ��ɫ�Իƽ���(Percaflavescens)�����������нϴ�Ӱ��; Oppedal ��[2]�����˹����ڶ��^������εĴ�������(Salmosalar)�ļ������������ջ�����Ӱ������; Taylor��[3]�о�����, �����ںͽϸߵĹ�ǿ���������
��������(Oncorhynchus mykiss)���������ʺ�����ת����; Wang ��[4]�о�����, 320~1 150 lx �Ĺ�ǿ���������б��ʯ����(Epinephelus coioides)����ijɻ��ʡ�������ؼ��ض������ʡ�Ŀǰ, �йع�ɫ�������ڡ���ǿ 3 �����������ѭ��ˮ��ֳϵͳ�д��������������ܼ���ʳ���ۺ�Ӱ���о���δ�����������о�����̽�����ڹ�ҵ��ѭ��ˮ��ֳģʽ��, ��ɫ�������ڡ���ǿ�� 3 ��������ӶԴ��������������ܼ���ʳ��Ӱ��, ּ�ڻ�ô���������������ʳ�����˹�ɫ�������ں�ǿ,��Ϊ LED ���ռ����ڴ�������ѭ��ˮ��ֳϵͳ�е�Ӧ���ṩ���۲ο���
����1 �����뷽��
����1.1 ����
��������������ɽ����������Ƽ��ɷݹ�˾�����о������ṩ, ʵ��ʱѡ���峤(39.71��1.58) cm,������(850.97�� 82.77) g �Ľ�׳�㡣ʵ��ѡ��Ų�����ʿ�����(Skretting)�����������, ����ҪӪ���ɷ�Ϊ: �ֵ��ס�48.0%, ��֬����18.0%, ����ά��1.0%, �ֻҷ֡�12.0%��ʵ��ϵͳ: ����������Ƶ� 3 ��ѭ��ˮ��ֳʵ��ϵͳ, ÿ��ϵͳ 3 ����ֳ��, ����ֳ�ء���Һ���������������ˡ���ĭ�����������������װ�á�����������ȵ�Ԫ���(ͼ 1)����ֳ��ΪԲ�β����ֽṹ, ֱ�� 200 cm, �ظ� 130 cm, ��Чˮ��Ϊ 105 cm��ÿ����ֳ�ص�ˮ�����Ϊ 3 297 L,��ˮ�سرڲ�������, ��Һ�������Ƽ����Ϳ��ƽ�ˮ��, ͨ�������ĵ����۹ܽ�����ˮ��ʵ����ð�ɫ(l550 nm)����ɫ(l455 nm)�ͺ�ɫ(l625 nm)�� COB ���ɷ�װ LED ��Դ���ƾ����й���ѧԺ�뵼���о������, �ɽ����������������Ƽ�����˾���졣��ǿ������ˮƽ��������� 1 ��ʾ�� ��ǿ��ˮ������λ���Ϸ� 5 cm �IJⶨֵΪ, ����Ͷ��Ƶĸ߶��Ա�֤����ʵ����Ĺ�ǿ��ʵ���ڼ�, �ڹⲼ������������ֳ��֮��, �Է�ֹ��ͬ��Դ����ߵ����Ⱦ����ɫ�����ǿ���ɺ���Զ�������Ϣ�ɷ�����˾���Ƶ� SP-10 �����Dzⶨ, �����ڲ���ʱ������������ڡ�


����1.2 ����
����1.2.1 ʵ���������� ʵ���� 2012 �� 8 ����2013 �� 3 �½���, Ϊ�� 180 d�� ʵ����� L9 (33) �������, ���ɫ�� �����ں�ǿ 3 ������, ÿ�������� 3 ��ˮƽ(�� 1), ���� 9 ���������: A1B1C1��A1B2C2�� A1B3C3�� A2B1C2�� A2B2C3�� A2B3C1��A3B1C3��A3B2C1��A3B3C2��ÿ�����������ֳ 60β��(���� PER ���ױ�־��� 30 β, δ��� 30 β),���� 540 β��ʵ���ڼ�, ÿ��7: 30 �� 14: 00 ��Ͷι�����������, Ͷ�����Ա�ʳΪ��ÿ��Ͷ��ʱ, �۲���Ⱥ��ʳ����Ծʱ��ֹͣͶι��ÿ��Ͷ�� 40 min ��, �ڹ�Һ���������۹ܴ�����ɴ����ռ��ж��ͷ��, ��¼δ��ʳ�Ķ�����, �������ʵ����ʳ����ʵ���, ˮ�±�����(15.7��0.4)�� , �� �� 24~26, pH7.2~7.5, �� �� �� �� �� �� Ϊ100%~ 140%, �ܰ��� (TAN)<0.25 mg/L����ˮ�����ݲⶨ��ˮ�����ݽ��е���, һ�㻻ˮ�������� 10%��1.2.2 ��Ʒ�ɼ� ʵ�鿪ʼ��, ÿ�½��㼢�� 1 d,����ÿ���������б�Ǻ�δ��ǵĴ����������������峤���泤, ͳ������Ͷι�����ж���, �ⶨ��������ʳ���ܡ�ÿ�¿ո��ɼ������ǵĴ�������ѪҺ, �Ʊ�Ѫ��, �����������ء� ��������ʱ,ͳ��ÿ��������ijɻ��ʡ�
����1.2.3 �������㹫ʽ
�����ɻ���(survival rate, SR): SR (%)=100��Nf/Ni[5]
�����������ض�������(specific growth rate, SGR):SGR (%/d)=100��(lnBW2�ClnBW1)/(T2�CT1)[6]�峤�ض�������(SGRBL): SGRBL (%/d)=100��(lnBL2�ClnBL1)/(T2�CT1)[6]������(daily weight gain, DWG): DWG (g/d)=(BW2�CBW1)/(T2�CT1)[6]�������(relative weight gain, RWG): RWG��(%)=100��BW2/BW1[7]������(condition factor, CF): CF=BW/FL3[7]
��������������ϵ��(coefficient of size variation,SV): SV=100��SD/X[6]
������ʳ��(food intake, FI): FI (%/d)=100��F/[0.5��(BW2+BW1)��(T2�CT1)][6]
��������ת��Ч��(food conversion efficiency, FCE):FCE (%)=100��(BW2�CBW1)/F[5]
��������ϵ��(food conversion ratio, FCR): FCR=F/(BW2�CBW1)[6]ʽ��, Ni��Nf �ֱ�Ϊ��ʼ��β����������β��;BW1��BW2 �ֱ�Ϊÿβ��ʵ���ʼ������������������(g); T1��T2�ֱ�ΪBW1��BW2����Ӧʱ��;BL1��BL2 �ֱ�Ϊ��ij�ʼ�峤�������峤(cm);BW Ϊÿβ���������(g); FL Ϊÿβ��泤(cm);F Ϊ����ʳ��(g); SD Ϊ����; X Ϊ���������ƽ��������(g)��
����1.2.4 �������صIJⶨ PER ���ױ�ǵ�ʵ������غ�, ����β����ȡѪ, Ѫ����ע����������еĺ������ƿ��������Ĺ���, Ȼ����5 000 r/min����10 min, ȡѪ���ڨC70���±���, ������������(GH)���������� Björnsson �� [8]�����ķ������߷����Ѫ������������(GH)��Ũ��, GH�������߷����Լ����ɱ����������\���о����ṩ��
����1.3 ͳ�Ʒ���
����ʵ�����ݲ��� SPSS18.0 ��������ͳ�Ʒ���,���õ����ط������(One-way ANOVA), ����С�������(LSD)�Ƚ����ݼ������ԡ����ݽ����ƽ��ֵ������( x ��SD)��ʾ, P<0.05 ʱ����������
����2 ��������
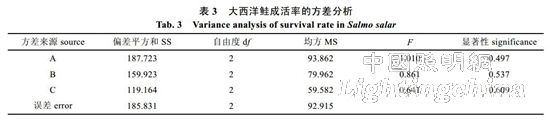
����2.1 ��ɫ�������ں�ǿ�Դ������ٳɻ��ʵ�Ӱ�촦���� 6 �� 9 �Ĵ������ٳɻ��ʾ�����������������(P<0.05, �� 2), ����� 8L�U16D �� 0.88W/m2 �� 4.55 W/m2 ���������������ѭ��ˮϵͳ�д������ٴ��� 9 ��ʵ�������з���ͳ��,�γɴ������ٳɻ����ɸߵ��͵Ĺ�ɫ����Ϊ: ��(95.00%)�����(90.55%)������(83.89%); ��������������Ϊ: 12L�U12D(93.33%)��24L�U0D(92.22%)��8L�U16D(83.89%); ��ǿ����Ϊ: 8.60 W/m2(94.44%)��0.88 W/m2(89.44%)��4.55 W/m2 (85.55%)�����, ���������ڰ⡢12L�U12D��8.60 W/m2 �������³ɻ�����ߡ�A���صİ�ͺ�������������,����A����ѡ����, ����ȷ����⡢12L�U12D��8.60 W/m2 Ϊ����������ֳ�����������������������ʾ(�� 3), ��ɫ�������ں�ǿ�Դ������ٳɻ��ʲ��첻����(P>0.05)�������Ϊ, �ڴ�����������������, ��ɫ�������ں�ǿ��δ�Դ������ٳɻ��ʲ���������Ӱ�졣

����2.2��ɫ�������ں�ǿ�Դ�������������Ӱ���ɱ�4�ɼ�,���������������ض������ʵı仯��ΧΪ0.18~0.71%/d, �ʲ�����ġ�W�������ơ�ʵ���30��60��90��120��150��, �����������ض������ʲ��첻����(P>0.05); ����180��ʱ,1��2��4��7��8�����첻����(P>0.05), ������������6��(P<0.05), �����������ض��������ɸ���������Ϊ: 1�顢4�顢2�顢7�顢8�顢3�顢5�顢9�顢6�顣�ɱ�5��֪, �峤�ض������ʱ仯������0.01~0.20%/d ֮��, ʵ���30��60��90��150��180��, �����峤�ض������ʲ��첻����(P>0.05), ���ڵ�120��ʱ2��5��6����������1��(P<0.05)���ӱ�6�ͱ�7����, ����ʵ��θ�����ķ����Ⱥ�������ؾ�����������(P>0.05), �仯��Χ�ֱ���1.08~1.31 �� 104.98%~130.00% ֮�䡣�ɱ�8�ó�, �����صı仯����Ϊ2.07~11.91g/d, ������������ر仯�������������ض�����������, ����30��60��90��120��150��, ������������ز��첻����(P>0.05), �ڵ�180��1��2��3��4��7��8��9����������6��(P<0.05), �����������ɸ���������Ϊ: 4�顢1�顢2�顢7�顢8�顢9�顢3�顢5�顢6�顣�ӱ�9��֪, ʵ��ǰ��������������ϵ�����첻����(P>0.05); ʵ����30��, 3����������1��4��8��(P<0.05), ��5��6��9����������4��(P<0.05); ����60��90�������첻���� (P>0.05); ��120 ��ʱ6 ���������� 8��(P<0.05); ����150��, 6��9����������7��(P<0.05);��180��, 9����������7��(P<0.05), ��������������ϵ���ɵ���������Ϊ9�顢5�顢6�顢4�顢2�顢3�顢1�顢8�顢7�顣��ͬ���������´�������Ѫ�����������غ�����ʱ�����Ƴʲ���ʽ�仯(��10)��ʵ�鿪ʼʱ��ʵ���30��,������Ѫ���е���������Ũ�Ⱦ��������Բ���(P>0.05); ��ʵ���60��, 3����������5��6��9��(P<0.05); ����90��, 8����������1��3��4��5��6��7 ��9��(P<0.05), ��2����������1��4��6 �� 7 ��(P<0.05); ����120��, 1��2��5 ��8����������6��(P<0.05), 5����������3��(P<0.05); ����150 ��, 2��3��4��9����������5��(P<0.05), 8����������1��5��6��7��(P<0.05), 9����������7��(P<0.05); ����180 ��, 9����������1��2��3��4��6��7 ��8��(P<0.05), ����9���������غ������(0.52 ng/mL)(�� 10)��







����2.3 ��ɫ�������ں�ǿ�Դ���������ʳ��Ӱ��ʵ��������, ��ǿ�Դ���������ʳ��(FI)Ӱ�����, ����ǹ����ڡ���ɫ, ������ΪC3B2A3, �� 8.60 W/m2��12L�U12D�����(�� 11)����ɫ������ת��Ч��(FCE)Ӱ�����, ����ǹ�ǿ������, ������Ϊ A1C3B1, ���⡢8.60W/m2��24L�U0D; A��B �����е�A1��A3��B1��B2��ˮƽ����������, ����A��B ���طֱ�ȡA3��B2, ����ȷ�������������A3C3B2, ����⡢8.60 W/m2��12L�U12D�� ��ɫ������ϵ��(FCR)Ӱ�����, ����ǹ����ں�ǿ, ���ˮƽ���Ϊ A2B3C2,�����⡢8L�U 16D��4.55 W/m2������ȷ�����������Ϊ��⡢12L�U12D��8.60 W/m2���Ա�11 ���ݵķ�������� F �������(�� 12), �ڱ�ʵ��������, ��ɫ�������ں�ǿ���Դ���������ʳ�ʡ�����ת��Ч�ʼ�����ϵ��û������Ӱ��(P>0.05)��
����3 ����
����3.1 ��ɫ�������ں�ǿ�Դ������ٳɻ��ʵ�Ӱ���ɫ�������ں�ǿ��Ӱ��Ӳ������ɻ��ʵ���Ҫ�������ӡ�Downing[9]�����˹��׳ɷֶԺ�����(Melanogrammus aeglefinus)�ɻ��ʵ�Ӱ�졣Trotter ��[10]�о�����, �ϳ��������ڿ�������������(Latris lineata)�ɻ���, ��24L�U0D �Ϻ��кڰ��ڵĹ����ڳɻ��ʽϸߡ�Puvanendran ��[11]�о�����, �� 2 400 lx ��ǿ�´�������(Gadusmorhua)�ɻ�����������300��600��1200 lx�顣��ʵ��������, �ں�⡢12L�U12D��8.60 W/m2��������, �������ٳɻ�����ߡ������ŵ�[12]���о���ʾ, ��(Hypophthalmichthys molitrix)����(Aristichthys nobilis)������10~12 hʱ�ɻ�����ߡ�Yoseda ��[13]���о�����, �ڸ߹�ǿ3 000��1000 lx ��, ���ǰ�(Plectropomus leopardus)�ɻ����������ڵ�ǿ(0 �� 500 lx)�顣��Щ�۵��뱾ʵ������һ��, �������������, ������12L�U12D���ϸ߹�ǿ(8.60 W/m2)������ߴ������ٳɻ��ʡ�
����

����3.2 ��ɫ�������ں�ǿ�Դ�������������Ӱ�����о�����, �����ڡ���ǿ���׳ɷ�(��ɫ)��Ӱ��Ӳ������������Ӧ����������Ϊ����Ҫ����[14�C16]���ڱ�ʵ����, ѡ�����������峤�ض������ʡ�����������ϵ���������غ�Ѫ�����������ص���Ϊ��ӳ������������ָ�ꡣ�ٿ�������,�������ص���Ҫ��������������л����ƽ����������[17]����ʵ������ʾ, ����ʱ������,�����ɫ�������ں�ǿ�Դ��������������ض������ʡ��峤�ض������ʡ�����������ϵ����������Ӱ����ֲ�ͬ�̶ȵIJ���, Ѫ�����������غ����ڵ� 180 ��ʱ������9(��⡢8L�U16D��4.55 W/m2)��������1��2��3��4��6��7 �� 8 �顣�����о�����, Karakatsouli ��[18]����(Cyprinuscarpio)���о�����, ��⡢�������, �����������شٽ��������ض������ʺ����������ӡ�Endal ��[19]���о���ʾ, �������ڻ�������տ�����ߴ��������������ٵ��������ܡ�Stefansson��[20]���о�����, ����Ȼ���������, �������տ��Դٽ�������������, ���������Ź�ǿ���߶����ӡ�Oppedal ��[21]���о���ʾ, �߹�ǿ�����ٽ��������д������ٵ�������Ҳ���о�ָ��, �����ڶ����������ÿ�����ͨ������−�����ᡱ�̼����������������صĺϳɺͷ��ڶ�����[22]��
����3.3��ɫ�������ں�ǿ�Դ���������ʳ��Ӱ���ɫ�������ں�ǿ��Ӱ��������ʳ����Ҫ������ Luchiari ��[23]���о�����, �����������������(Sander lucioperca)�������ʳ�ʺ�����Ч�ʡ�Biswas ��[24]������(Pagrus major)���о���ʾ,������ 24L�U0D �µ���ʳ�ʺ�����ת��Ч����������16L�U8D��6L�U6D��12L�U12D��Strand ��[25]���о�����, ��ǿ������Ӱ��ŷ����(Perca fluviatilis)����ʳ�ʡ��ڱ�ʵ����, ������ӶԴ���������ʳ�ʡ�����ת��Ч�ʼ�����ϵ����Ӱ���ɸߵ�������Ϊ: ��ǿ�������ڡ���ɫ, ��ɫ����ǿ�������ڼ���ɫ�������ڡ���ǿ, ������Ͼ�Ϊ8.60 W/m2��12L�U12D����⡣��������������,��ɫ�������ں�ǿ��δ�Դ���������ʳ�ʡ�����ת��Ч�ʼ�����ϵ������������Ӱ�졣��ʵ������ǰ�����ı�������һЩ����, ���ֲ����ԭ��������һ���о���
����4 ����
�����ڹ�ҵ��ѭ��ˮ��ֳϵͳ��, ��ɫ�������ں�ǿ�Դ���������������ʳӰ�첻����, ��⡢12L�U12D��8.60 W/m2 �DZ�ʵ�������½�Ϊ���˵Ĺ���������LED ����Ϊһ�����ͽ��ܹ�Դ, ���ڵ���ѭ��ˮ��ֳϵͳ���������������ʳ�нϺõ�Ӧ��ǰ����
�����ο����ף�
����[1] Head A B, Malison J A. Effects of lighting spectrum anddisturbance level on the growth and stress responses of yellow perch Perca flavescens[J]. J World Aquacult Soc, 2000,31 (1): 73�C80.
����[2] Oppedal F, Berg A, Olsen R E, et al. Photoperiod in seawaterinfluence seasonal growth and chemical composition in autumn sea transferred Atlantic salmon (Salmo salar L.) giventwo vaccines[J]. Aquaculture, 2006, 254 (1�C4): 396�C 410.
����[3] Taylor J F, North B P, Porter M J R, et al. Photoperiod canbe used to enhance growth and improve feeding efficiency infarmed rainbow trout, Oncorhynchus mykiss[J]. Aquaculture,2006, 256 (1�C4): 216�C234.
����[4] Wang T, Cheng Y Z, Liu Z P, et al. Effects of light intensityon growth, immune response, plasma cortisol and fatty acidcomposition of juvenile Epinephelus coioides reared in artificial seawater[J]. Aquaculture, 2013, 414: 135�C139.
����[5] ����, ��С��, �����, ��. ���� Vc �Բ�������������Ʒ�ʼ������������ߵ�Ӱ��[J]. �й�ˮ����ѧ, 2008, 15 (5):794�C800.
����[6] �Ž���, ���ظ�, ����. �л������������ӵ��в�ȵ���������ʳ����Ϊ��Ӧ[J]. �й�ˮ����ѧ, 2013, 20(3):592�C598.
����[7] Leclercq E, Taylor J F, Sprague M, et al. The potential ofalternative lighting-systems to suppress pre- harvest sexualmaturation of 1+ Atlantic salmon (Salmo salar) post-smoltsreared in commercial sea-cages[J]. Aquaculture, 2011, 44 (2):35�C47.
����[8] Björnsson B T, Taranger G L, Hansen T, et al. The interrelation between photoperiod, growth hormone and sexualmaturation of adult Atlantic salmon (Salmo salar)[J]. GenComp Endocrinol, 1994, 93 (1): 70�C81.
����[9] Downing G. Impact of spectral composition on larval haddock, Melanogrammus aeglefinus L., growth and survival[J].Aquacult Res, 2002, 33 (4): 251�C259.
����[10] Trotter A J, Battaglene S C, Pankhurst P M. Effects ofphotoperiod and light intensity on initial swim bladderinflation, growth and post-inflation viability in culturedstriped trumpeter (Latris lineata) larvae[J]. Aquaculture,2003, 224 (1�C4): 141�C158.
����[11] Puvanendran V, Brown J A. Foraging, growth and survivalof Atlantic cod, Gadus morhua, larvae reared in differentlight intensities and photoperiods[J]. Aquaculture, 2002, 214��(1�C4): 131�C151.
����[12] ������, �Ե���, �ž�ȫ. ��ͬ����ʱ�������㡢�����������ʹ���ʵ�Ӱ��[J]. ��̬ѧ��־, 1994, 13(3):41�C44.
����[13] Yoseda K, Yamamoto K, Asami K, et al. Influence of lightintensity on feeding, growth, and early survival of leopardcoral grouper (Plectropomus leopardus) larvae undermass-scale rearing conditions[J]. Aquaculture, 2008, 279(1�C4): 55�C62.
����[14] Oppedal F, Taranger G L, Juell J E, et al. Growth, osmoregulation and sexual maturation of underyearling Atlanticsalmon smolt Salmo salar L. exposed to different intensitiesof continuous light in sea cages[J]. Aquacult Res, 1999, 30(7): 491�C499.
����[15] Boeuf G, Le Bail P Y. Does light have an influence on fishgrowth?[J]. Aquaculture, 1999, 177 (1�C4): 129�C152.
����[16] Karakatsouli N, Papoutsoglou S E, Pizzonia G, et al. Effectsof light spectrum on growth and physiological status of gilthead seabream Sparus aurata and rainbow trout Oncorhynchus mykiss reared under recirculating system conditions[J].Aquacult Eng, 2007, 36 (3): 302�C309.
����[17] Björnsson B T. The biology of salmon growth hormone:from daylight to dominance[J]. Fish Physiol Biochem, 1997,17 (1/6): 9�C24.
����[18] Karakatsouli N, Papoutsoglou S E, Sotiropolos N, et al. Effects of light spectrum, rearing density and light intensity ongrowth performance of scaled and mirror common carp Cyprinus carpio reared under recirculating system conditions[J].Aquacult Eng, 2010, 42 (3): 121�C127.
����[19] Endal H P, Taranger G L, Stefansson S O, et al. Effects ofcontinuous additional light on growth and sexual maturity inAtlantic salmon, Salmo salar, reared in sea cages[J]. Aquaculture, 2000, 191 (4): 337�C349.
����[20] Stefansson S O, Hansen T, Taranger G L. Growth andparr-smolt transformation of Atlantic salmon (Salmo salar)under different light intensities and subsequent survival andgrowth in seawater[J]. Aquacult Eng, 1993, 12 (4): 231�C243.
����[21] Oppedal F, Taranger G L, Juell J, et al. Light intensity affects growth and sexual maturation of Atlantic salmon(Salmo salar L.) postsmolts in sea cages[J]. Aquat LivingRes, 1997, 10: 351�C357.
����[22] Björnsson B T, Thorarensen H, Hirano T, et al. Photoperiodand temperature affect plasma growth hormone levels,growth, condition factor and hypoosmoregulatory ability of juvenile Atlantic salmon (Salmo salar) during parr-smolttransformation[J]. Aquacultue, 1989, 82 (1�C4): 77�C91.
����[23] Luchiari A C, Freire F A M, Pirhonen J, et al. Longer wavelengths of light improve the growth, intake and feed efficiency of individually reared juvenile pike perch Sanderlucioperca (L.)[J]. Aquacult Res, 2009, 40 (8): 880�C886.
����[24] Biswas A K, Seoka M, Inoue Y, et al. Photoperiod influencesthe growth, food intake, feed efficiency and digestibility ofred sea bream (Pagrus major)[J]. Aquaculture, 2005, 250 (3�C4): 666�C673.
����[25] Strand Å, Alanärä A, Staffan F, et al. Effects of tank colourand light intensity on feed intake, growth rate and energyexpenditure of juvenile Eurasian perch, Perca fluviatilis L.[J]. Aquaculture, 2007, 272 (1�C4): 312�C318.Effects of different types of environment light on the growth performance and feeding of Atlantic salmon (Salmo salar) in recirculatingaquaculture systemsQIU Denggao1, 2, 3, XU Shihong1, LIU Ying1, SONG Changbin4, CHI Liang1, WANG Shunkui5, YU Kaisong5
����1. Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China;
����2. University of Chinese Academy of Sciences, Beijing 100049, China;
����3. Fisheries Research Institute of Fujian, Xiamen 361012, China;
����4. Institute of Semiconductors, Chinese Academy of Sciences, Beijing 100083, China;
����5. Shandong Oriental Ocean Sci-Tech Co. Ltd., Yantai 264003, China
����Abstract: We aimed to explore the effects of different types of light color(i.e., A1, white light; A2, blue light; A3, red light), photoperiod (i.e., B1, 24L�U0D; B2, 12L�U12D; B3, 8L�U16D) and light intensity (i.e., C1, 0.88 W/m2; C2, 4.55 W/m2; C3, 8.60 W/m2) on the growth performance and feeding of Atlantic salmon (Salmo salar) with initial body weights of 850.97��82.77 g for 180 days in recirculating aquaculture systems (RAS) using orthogonal test methods.Nine treatment groups were designed in the present experiment: A1B1C1(group1), A1B2C2(group2), A1B3C3(group3),A3B3C2(Group4), A2B2C3(group5), A2B3C1(group6), A3B1C3(group7), A3B2C1(group8), and A1B1C1(group9). The results showed that the highest survival rate was found in red light, 12L�U12D and 8.60 W/m2 at the end of the trial, andno significant difference was observed in the survival rates of the experimental groups (P>0.05). There were no significant differences in relative weight gain and condition among the nine groups at each sampling time (P>0.05). At day120, the specific growth rate of body length for groups 2, 5 and 6 were significantly higher than those of group 1(P<0.05). At day 180, the specific growth rate of body weight in groups 1, 2, 4, 7 and 8 were significantly higher thanthose of group 6 (P<0.05), and the daily weight gain of groups 1, 2, 3, 4, 7, 8 and 9 showed higher variation than thoseof group 6 (P<0.05); the coefficient of size variation (SV) in group 9 was lower than in group 7 (P<0.05). Additionally,the plasma growth hormone in group 9 was a higher concentration than that of groups 1, 2, 3, 4, 6, 7 and 8 (P<0.05).Better food intake(FI), food conversion efficiency(FCE) and food conversion ratio(FCR) were also observed in red light(12L�U12D) at 8.60 W/m2 at the end of the experiment, while light color, photoperiod and light intensity had no significant effects on the FI, FCE and FCR between groups(P>0.05). Hence, red light (1




�й�����������Ƶ��������ҵ����������������������Ҫ����м����ԡ��ɶ��ԡ���ӭ�о�������ԺУ����ҵ����Ͷ�塣
Ͷ�����䣺edit@lightingchina.com.cn
��ϵ�绰��0086-020-85530605-5029
(Ͷ��ʱ��ע��������������λ���ʱ�͵�ַ���绰��E-mail���Ա�֪ͨ��˽�����緢����������֪ͨ�������ѯ��)
�㶫��������ý����˾ ��Ȩ���С���ֵ����ҵ��Ӫ����֤����B2-20050039����ICP��06007496��
���棺020-85548112��E-mail��Service@lightingchina.com.cn �й�������



